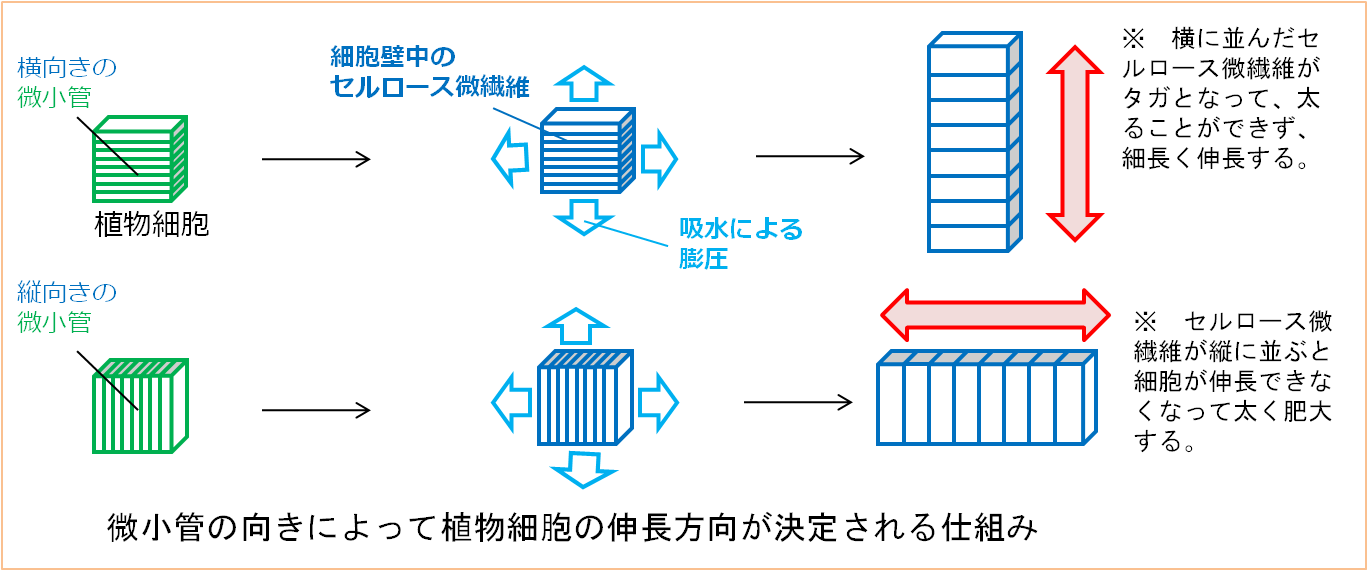
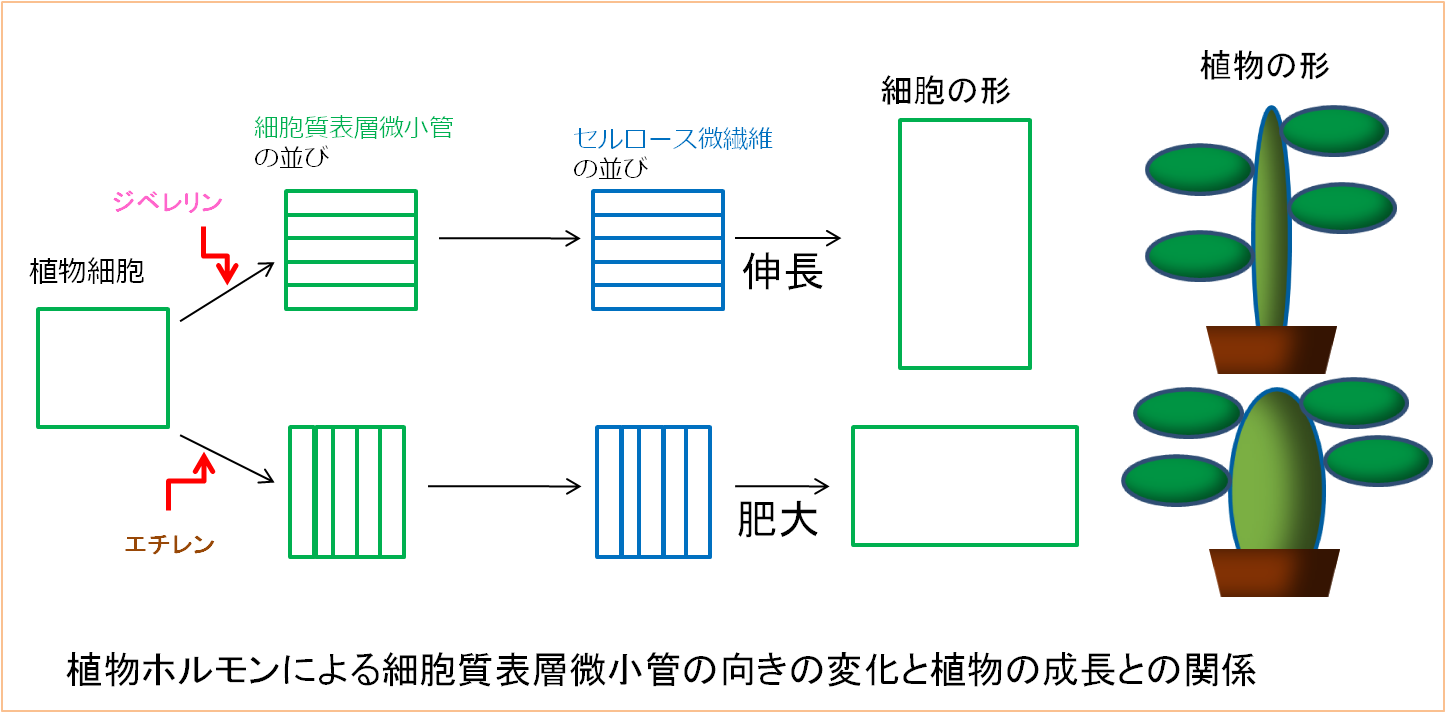
被子植物では、細胞質表層微小管の向きは植物ホルモンによって制御されており、細胞質表層微小管の配向の制御は植物ホルモンによる植物体の成長制御において重要な仕組みのひとつです。
植物ホルモンの一種であるジベレリンは、植物の茎を細長く伸長させる働きをもっています。このジベレリンによる茎の伸長促進作用は、茎の細胞の細胞質表層微小管の向きを伸長方向に対して垂直に変化させることによるものであることが判っていますが、その向きが変化する仕組みについては明らかにされていません。私たちの研究室では、主にアズキの芽生えの茎切片を利用した実験系により、このジベレリンによる細胞質表層微小管の配向変化機構を解明する目的で研究を行っています。
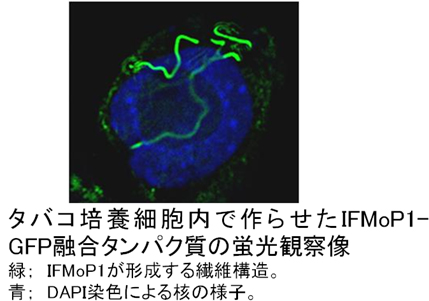
そのために、ジベレリンによって発現誘導される遺伝子の発現や機能、植物ホルモンを作用させた時の微小管の安定性の変化やアクチンフィラメントという微小管とは別のタンパク質繊維の役割などに着目した研究を行っています。蛍光タンパク質によって標識したチューブリン(微小管の構成タンパク質)を導入した形質転換タバコ培養細胞による、細胞質表層微小管の動きの観察も行っています。